LIMBO: Analysis of
hippocampal
circuitry
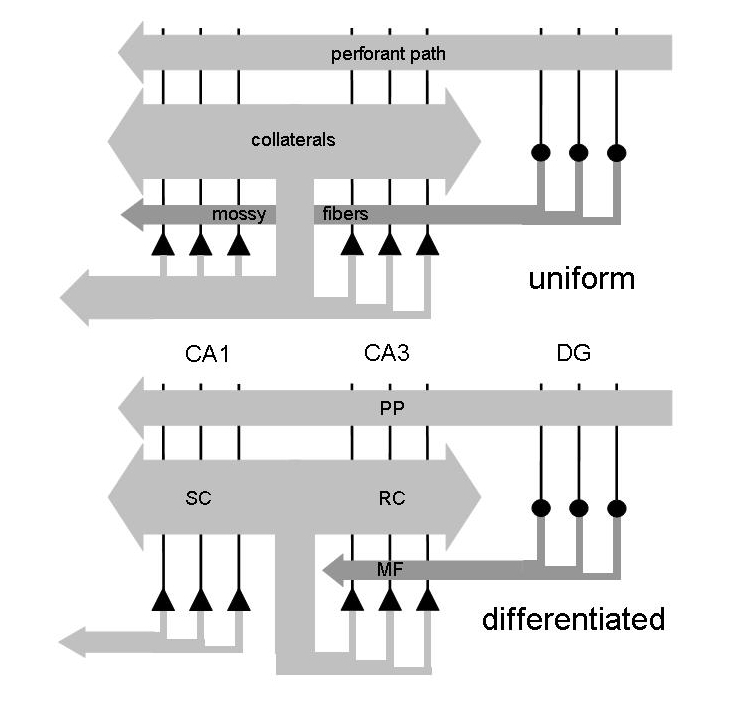
The aim
is to understand the relation between the organization
of the
hippocampus
and its function in memory formation [2,4], along the lines suggested
by
David Marr. The central hypothesis, shared with several investigators,
is that CA3 can be viewed as the core autoassociator [5]: we have shown
that even with the storage of multiple spatial maps essentially the
same
constraints apply as with discrete "episodic memory" firing patterns
[9].
Our early proposal of a specific role for each of the two main
input
systems to the CA3 field [1] is consistent with experimental findings
[see JM
Lassalle, T Bataille & H Halley, Neurobiol
Learning
and
Mem 73:243-257 (2000) and I Lee & RP
Kesner, Hippocampus 14:66-76 (2004)].
An analytical approach has been introduced for understanding
quantitatively the operations performed by the
Schaffer
collaterals [3,7], and later extended to include the effect of direct
entorhinal
projections to CA1 [8]. Similar measures have been applied, in
collaboration with Edmund Rolls and his lab, to monkey hippocampal
recordings [6], and to rat hippocampal multi-unit recordings
from the lab of Carol Barnes and Bruce McNaughton.
An approach based on simulations
has been introduced more recently [11], modeled in part after
neurophysiological recordings from the
Tucson
lab, and applied to assess the neural mechanisms that may underlie
temporal
prediction
in the hippocampus: associative plasticity and firing rate adaptation.
The simulations offer a general quantitative approach to address the
main outstanding question [3,10]: what
is the exact contribution of CA1 to hippocampal processing? or, in
other words, what is the computational significance of the CA3-CA1
differentiation in mammals [12]? A new light on this issue has been
cast by exciting novel findings from the Trondheim lab
[13], with whom is active an intense collaboration,
and whose website you should
visit for more
information on their experiments [14]. A wider collaboration has started since February, 2008 within the EU SPACEBRAIN project.
The
discovery of grid cells in Trondheim, and of their locally coherent
transition dynamics when CA3 cells undergo global remapping [15] has
stimulated ideas about the origin of the grid fields [16] which
contrast with the popular attractor and interfering oscillator models;
and it has also led to reassess the role of the dentate gyrus
[17] in
generating new CA3 representations (current work by EC, AT). The
theoretical context for this work and its commonalities to seemingly
unrelated research is reviewed in [18].
References:
- AT & ET Rolls, Hippocampus
2:189-199 (1992). Poorly scanned copy.
- AT & ET Rolls, Hippocampus
4:374-391 (1994). Poorly scanned copy.
- AT, J Comput Neurosci 2:259-272
(1995)
- AT, WE Skaggs & CA Barnes,
Hippocampus
6:666-674 (1996) You are welcome to ftp an early draft.ps
- ET Rolls, AT, D Foster & C
Perez-Vicente,
Neural Networks 10:1559-1569 (1997)
- ET Rolls, AT, RG Robertson, P
Georges-Francois
& SP, J
Neurophysiol 79:1797-1813
(1998)
- SRS, SP, ET Rolls & AT, proceedings
in Information Theory and the Brain, p. 257-272, Cambridge UP, (2000)
- CFM, SP, ET Rolls & AT, proceedings
in Information Theory and the Brain, p. 273-289, Cambridge UP, (2000)
- FPB & AT, Phys Rev E 58:7738
(1998)
- AT & IS, Cognitive
Neuropsychology
12:557-575
(2002). You are welcome to download a draft.ps
- AT, Hippocampus 14:539-556 (2004).
You are welcome to download a draft.pdf
- AT, Neuroinformatics 2:361-366
(2004). You are welcome to download a draft.pdf
- S
Leutgeb, JK Leutgeb, AT, M-B Moser & EI Moser,
Science 305:1295-1298 (2004)
- JK Leutgeb, S Leutgeb, AT, R Meyer, CA Barnes,
BL McNaughton, M-B Moser & EI Moser, Neuron 48:345-358 (2005)
- M Fyhn, T Hafting, AT, MB Moser, EI Moser, Nature 446: 190-194 (2007)
- EK, AT, manuscript submitted to Hippocampus (2008)
- AT, A Tashiro, MP Witter, EI Moser, Neuroscience 154:1155-1172 (2008)
- AT, Ch.3 in Cognitive Biology (Tommasi et al, eds), MIT press (2009)
Last updated 06/07/08. Back to LIMBO,
CNS,
SISSA.